- Половые клетки
- (синоним
гаметы) — специализированные клетки, обладающие гаплоидным набором
хромосом и обеспечивающие при половом размножении передачу наследственной
информации от родителей к потомству
.
Половые клетки происходят от диплоидных клеток в
результате редукции числа хромосом при эволюции полового процесса. У
большинства многоклеточных организмов первичные
половые клетки (гоноциты) обособляются
вследствие первых делений в начале эмбриогенеза из экто- или энтодермы, а затем
различными путями перемещаются в формирующиеся половые железы. У особей
мужского пола в половых железах образуются мужские
половые клетки , или сперматозоиды, а
у женского —яйцеклетки. Организм с истинным
гермафродитизмом продуцирует оба типа
половых клеток . Процесс развития
половых клеток рассматривается как начальный этап
онтогенеза
и называется гаметогенезом, образование сперматозоидов — сперматогенезом,
а яйцеклеток — оогенезом. Зрелые
половые клетки возникают в процессе
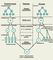
гаметогенеза, который условно подразделяют на 4 периода — размножение,
рост, созревание и формирование (
рис.
1). В периоде размножения диплоидные клетки сперматогонии и оогонии
(предшественники гамет) несколько раз делятся митотически, обеспечивая
нарастание числа будущих
половых клеток . Интенсивное размножение особенно характерно для
сперматогонии. В периоде роста происходит увеличение размеров клеток,
сопровождающееся накоплением в них питательных веществ, РНК, ряда структурных
белков. Значительный рост клеток ярко выражен в оогенезе. Основным содержанием
периода созревания является мейоз, в результате которого из каждой диплоидной
клетки-предшественницы образуется 4 клетки с гаплоидным набором хромосом. При
сперматогенезе эти клетки одинаковы по размеру и позже становятся
сперматозоидами, а при оогенезе мейоз обеспечивает неравномерное деление
цитоплазмы. В результате лишь одна гаплоидная клетка из четырех становится
яйцеклеткой, способной к оплодотворению, а три другие представляют собой
редукционные тельца, содержащие избыток хроматина и в конечном счете
погибающие. Кроме обеспечения гаплоидности мейоз приводит также к возникновению
качественного многообразия половых клеток. В профазе первого мейотического
деления гомологичные хромосомы отцовского и материнского происхождения,
спирализуясь, сближаются попарно соответствующими друг другу участками (так
называемая конъюгация), образуя биваленты. При этом отдельные хроматиды
переплетаются между собой и могут разрываться в аналогичных участках. В
процессе восстановления целости хроматид гомологичные хромосомы способны
обмениваться соответствующими участками. Этот процесс называется
кроссинговером. В анафазе первого мейотического деления происходит независимое
расхождение материнских и отцовских хромосом к полюсам
клетки,
вследствие
чего в гаплоидном наборе будущих гамет возникают разные сочетания материнских и
отцовских хромосом. Последний период гаметогенеза (период формирования)
наблюдается только при сперматогенезе, во время которого гаплоидные
клетки — сперматиды — приобретают особенности строения, характерные
для зрелых сперматозоидов. Сперматогенез у самцов млекопитающих протекает
постоянно на протяжении всего периода половой зрелости и мало зависит от
действия гормональных факторов. В отличие от этого этапы оогенеза не только
растянуты во времени, но и гормонально зависимы. Запас ооцитов, содержащихся
при рождении в яичниках самок, в дальнейшем не пополняется. Данная
закономерность присуща и гаметогенезу человека. Так, в яичнике новорожденной
девочки содержится около 2 млн. ооцитов, из них достигают зрелости и
претерпевают овуляцию только приблизительно 400. Первое мейотическое деление в
ооците происходит в момент овуляции, второе — уже после проникновения в
него сперматозоида.
Зрелая яйцеклетка имеет неоднородную
биохимическую организацию. На анимальном ее полюсе увеличена концентрация
гликогена и РНК, по экватору располагается аскорбиновая кислота, желток
собирается в основном на вегетативном полюсе. Такая неоднородность цитоплазмы
называется ооплазматической сегрегацией. Она имеет большое значение для
детерминации и последующей дифференцировки зародыша
на стадиях дробления и гаструляции. На характер дробления влияют также
количество и распределение желтка в яйцеклетке. Снаружи яйцеклетка покрыта
кроме цитоплазматической мембраны также и дополнительными оболочками. Так,
яйцеклетка млекопитающих и человека имеет блестящую оболочку — zona
pellucida, окруженную фолликулярными клетками, участвующими в ее метаболизме (рис. 2). Для оплодотворения необходимо
сближение мужских и женских половых клеток , которое достигается благодаря осеменению. У
большинства водных животных осеменение наружное, у наземных животных —
внутреннее, при котором сперматозоиды попадают непосредственно в половые пути
самки. Встрече мужских и женских гамет (рис.
3) и предохранению яйцеклетки от внедрения в нее избыточного количества
сперматозоидов способствуют выделяющиеся П. к. специфические вещества —
гино- и андрогамоны. В момент контакта половых клеток происходит выделение
ферментов акросомы сперматозоида и растворение участка яйцевой оболочки. При
этом объединяется содержимое обеих гамет и образуется единая клетка —
зигота. У некоторых организмов ядра сперматозоида и яйцеклетки (пронуклеусы)
сливаются вскоре после оплодотворения. У млекопитающих и человека их
объединение происходит в метафазе первого деления — дробления.
Биологическое значение оплодотворения
заключается в восстановлении в зиготе диплоидности хромосомного набора,
характерного для данного вида организмов, и в возникновении ее генетической
уникальности в результате объединения наследственного материала хромосом
родительских особей, претерпевшего предварительную рекомбинацию в процессе
мейоза. Кроме того, проникновение сперматозоида в яйцеклетку обеспечивает
активацию в ней различных синтетических процессов и начало дробления.
Библиогр.:
Газарян К.Г. и Белоусов Л.В. Биология индивидуального развития
животных, М., 1983; Карлсон Б. Основы эмбриологии по Пэттену, пер. с
англ., т. 1, М., 1983.